I’m not sure who is looking at only “one trait”. Here are some extracts from the paper ‘Genetic Diversity and Population Structure of Two Tomato Species from the Galapagos Islands’ - Yveline Pailles et al - 2017:
S. cheesmaniae and S. galapagense Can Be Clearly Differentiated by Genetic Analysis
To dissect the pattern of genetic variation among the accessions, we used both PCA from EIGENSOFT 6.0 (Price et al., 2006) and Bayesian clustering from STRUCTURE software (Pritchard et al., 2000). For PCA, the SNP data was further pruned based on LD, to obtain a subset of SNPs that are in approximate linkage equilibrium with each other. PLINK (Purcell et al., 2007) was used to calculate LD, based on pairwise genotypic correlation with a threshold of r 2 = 0.8. LD-based pruning reduced the SNP marker collection to 2,428 SNPs.
The PCA showed a clear division between the accessions identified as S. galapagense and S. cheesmaniae, as well as from S. lycopersicum and S. pimpinellifolium (Figure 3A).
The accessions belonging to S. galapagense cluster closely together (orange diamonds), while S. cheesmaniae accessions are also clustered (green circles), with the exception of two accessions: LA0531 and LA3124 (marked by an arrow), which could be admixtures. Individual PCAs of the accessions belonging to each of S. galapagense and S. cheesmaniae can be found in the Supplementary Materials (Supplementary Figure S2).
The groups obtained from the PCA were identical with those formed by an alternative clustering program, STRUCTURE (Pritchard et al., 2000), which uses a Bayesian clustering approach to identify the number of populations (K) with the highest structure (Supplementary Figure S3). This is determined by plotting Delta K, based on the method of Evanno et al. (2005); our DeltaK plot showed a peak at K = 3 (Supplementary Figure S4), suggesting the presence of three genetically distinct groups that differentiate the two Galapagos tomato species and the tomatoes native to mainland South America (Figure 3B). Once more, accessions LA0531 and LA3124 appear to be genetic admixtures. The admix nature of these two accessions was confirmed by the ancestry membership coefficients (Q), which show that LA0531 belongs to both cluster 1, formed by the rest of S. cheesmaniae accessions (Q = 0.674), and to cluster 2, formed by all S. galapagense accessions (Q = 0.325), whereas, LA3124 appears to be part of cluster 1 (Q = 0.432) and cluster 3, formed by S. lycopersicum and S. pimpinellifolium (Q = 0.568) (Supplementary Table S3). The collection notes of these two accessions, obtained from TGRC database 1 (Supplementary Table S4), report morphological differences from the typical S. cheesmaniae since the time of their collection (Supplementary Figure S1), which confirm that they are hybrids and crossing did not occur during later seed propagation, but in the natural environment. It is also worth noting, that the two accessions are unique to Baltra and Santa Fe, respectively, two very small islands – only one accession was collected from each island (Supplementary Table S1).
The results from PCA and STRUCTURE are largely consistent. For both analyses we found that S. galapagense, S. cheesmaniae, S. lycopersicum, and S. pimpinellifolium accessions are clearly separated. Although the sampling size of S. lycopersicum and S. pimpinellifolium accessions is small, the three reference sequences clustered together in all of the analyses performed. Thus, they provide useful reference points to facilitate estimation of the genetic distance between the Galapagos tomato populations.
A bit further into the paper:
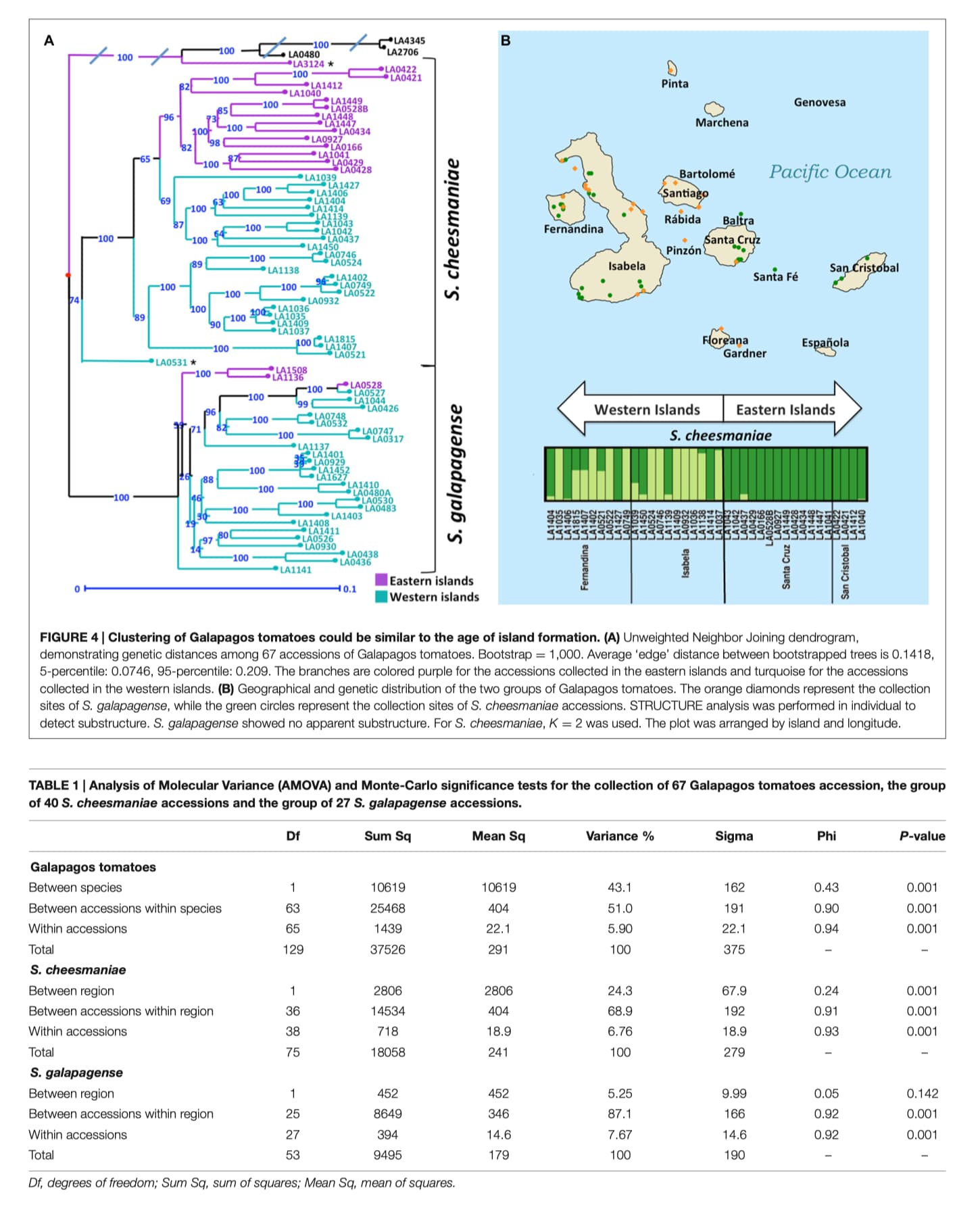
Clustering of S. cheesmaniae Accessions Could Be Similar to the Age of Island Formation
In the genetic distance tree, the S. cheesmaniae accessions separate into two sub-clusters with a 100% bootstrap support, while the admixtures remained separated from the main S. cheesmaniae branches (Figure 4A). Likewise, the population structure inferences using STRUCTURE (Figure 4B) of S. cheesmaniae accessions show two sub-clusters (K = 2) and no further structure within the S. galapagense group (K = 1) (Supplementary Figure S5). S. galapagense showed no structure, even when using the No admixture model in STRUCTURE (Pritchard et al., 2000; data not shown).
Interestingly, the division of the S. cheesmaniae cluster shows clear geographic structure. The grouping of the accessions matches their region of origin: the accessions collected in the western islands separate clearly from those collected in the eastern islands. This can be linked to the island formation timeline, as all the islands originated at the volcanic hotspot and then moved east with the Nazca plate which holds the Galapagos archipelago (Geist et al., 2014). The eastern islands are therefore older than the western islands (Geist et al., 2014) and from this we can infer that those populations found in the eastern islands may be older than the ones found in the western islands; alternatively, they could be ancestors of the accessions in the west. This inference is supported by the comparison of Wright’s fixation index (F ST ) values (Wright, 1951) for each S. cheesmaniae cluster, obtained from the STRUCTURE analysis (Pritchard et al., 2000). The mean F ST -value of the accessions from the western islands (0.6239), is considerably higher than the mean F ST -value of the accessions from the eastern islands (0.2790), which suggests the occurrence of a strong episode of genetic drift on those populations from the newer islands (Falush et al., 2003).
Analysis of Genetic Variation in Galapagos Tomatoes
An AMOVA (Excoffier et al., 1992) was performed to examine patterns of genetic variation and to estimate variance components at the levels of species and accessions. AMOVA showed that 43.1% of the total variation in the Galapagos tomato populations was explained by differences between the two species; whereas 51% was explained by differences between accessions within the species (Table 1). This confirms that the two species are considerably different, but also there is great variation among the accessions within a species.
Best to read the paper for full details but here’s another graphic from a bit further on:
I like this paper because speculation can be useful and interesting, but I do love a good evidence-based approach also. It would seem that from a scientific perspective, these two Galapagos species are clearly differentiated, and, clearly differentiated also from lycopersicum and pimpinellifolium.